Плацента человека относится к гемохориальному типу, который характеризуется непосредственным контактом материнской крови и хориона вследствие нарушения целости слизистой оболочки матки со вскрытием е сосудов. Возникнув на высшей ступени эволюции, гемохориальная плацента способствует наиболее полном осуществлению сложных морфофункциональных взаимоотношений — между матерью и плодом. Установлению контакта материнских и зародышевых тканей предшествуют дробление оплодотворенной яйцеклетки и превращение эндометрия в децидуальную ткань.
Вне беременности покровный эпителий слизистой оболочки матки содержит два типа клеток—секреторные и мерцательные, соотношение которых зависит от соотношений половых гормонов. В пред имплантационном периоде мерцательные клетки отсутствуют, а в секреторных клетках обнаруживаются признаки высокой активности. Наряду с микроворсинками в этих клетках выявляются цитоплазматические выросты, занимающие значительную часть апикальной поверхности. В них различимы митохондрии, везикулы, заполненные содержимым различной электронной плотности, частицы гликогена. Предполагают, что такие выросты служат проявлением секреции, а возможно, пиноцитарной активности покровного эпителия матки. После прикрепления бластоцист к эндометрию микроворсинки перестают быть видимыми, полностью исчезают липиды и снижается активность неспецифических эстераз.
Середина секреторной фазы характеризуется значительным утолщением стенок спиральных артерий, образующих мощные «клубки» в спонгиозном и компактном слоях (рис. 1).
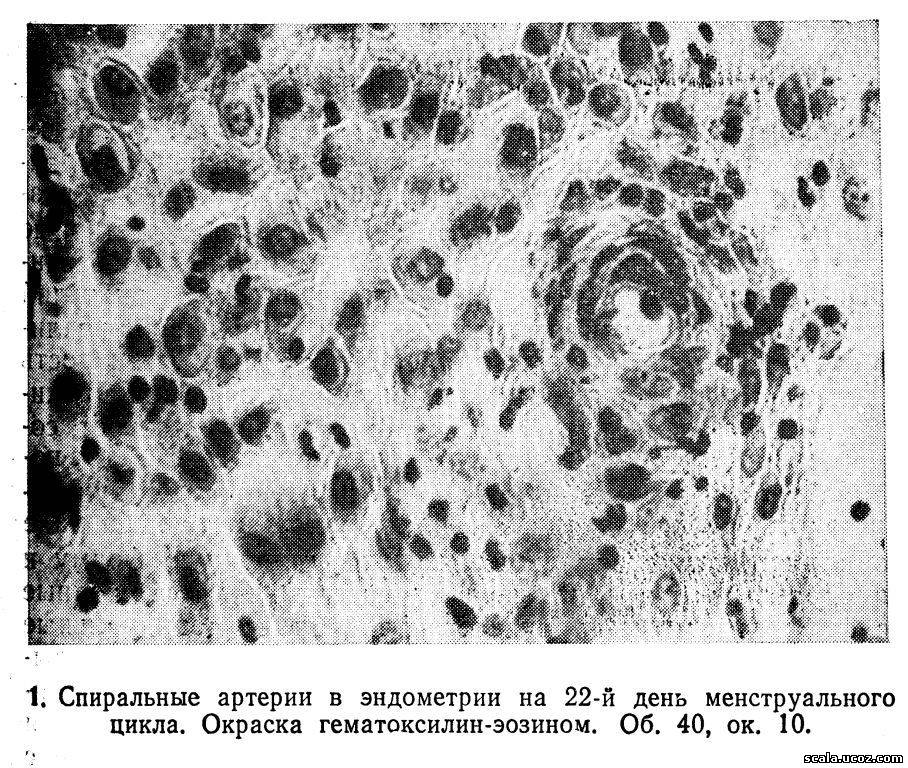
Развитие клубков спиральных артерий обусловлено воздействием прогестерона на эндометрий. В связи с изменением скорости кровотока и повышением проницаемости сосудистых стенок под влиянием половых гормонов развивается отек слизистой оболочки, достигающий максимума к 20-му дню цикла.
Дробящееся яйцо движется по яйцеводу в течение дней и оказывается в матке в стадии ранней бластоцисты. Бластомеры наружного слоя образуют внезародышевую эктодерму — трофобласт. Из бластомеров, находящихся внутри (эмбриобласт), формируются провизорные органы и зародышевый узелок, который накапливающейся жидкостью оттесняется к одному из полюсов бластоцисты. В полости матки бластоциста может находиться в свободном состоянии 3—4 суток. Имплантация происходит на 6—7-е сутки после оплодотворения. Утратив блестящую оболочку, бластоциста прикрепляется к эндометрию, как правило, в области зародышевого узелка, а через 2 суток оказывается целиком погруженной в слизистую оболочку матки и покрытой сверху «пробкой» фибрина. Процесс растворения блестящей оболочки протекает только в присутствии маточного секрета содержащего в-гликопротеиды в определенной концентрации. Противостояние бластоцисты и слипание трофобласта с поверхностью эпителия эндометрия относятся к основным стадиям имплантации, во время которых происходят морфофункциональные изменения бластоцисты и маточного эпителия.
Имплантация осуществляется на 20—21-й день нормального 28-дневного менструального цикла, т. е. в середине стадии секреции, когда функциональный слой эндометрия имеет наибольшую толщину. В нем четко различимы спонгиозный и компактный слой, а секреторные превращения эндометрия достигают наибольшего развития. Железы эндометрия расширены, извиты, эпителий желез с неровными апикальными краями содержит липиды, гликоген, кислую глицерофосфатазу. Активность щелочной фосфатазы слабо выражена или не обнаруживается. В просветах желез имеется слизистый секрет, содержащий мукоиды, глюкопротеиды, гликоген. В этот период клетки стромы эндометрия (фиброциты), окружающие сосуды компактного слоя, увеличиваются. Вначале они располагаются рыхло, имеют удлиненную форму и превращаются в молодые децидуальные клетки, сходные с фиброцитами. В процессе созревания децидуальные клетки значительно укрупняются и становятся округлыми или овальными, с крупным светлым ядром, тесно прилежат друг к другу. Пусковой механизм превращения эндометрия в истинную децидуальную ткань связан с выделением бластоцистой гистамина.
После имплантации бластоцисты железы компактного слоя постепенно суживаются, просвет их становится щелевидным (рис. 2), а с 8-й недели беременности почти неразличим. Количество децидуальных клеток компактного слоя значительно увеличивается, в первые недели беременности в них наблюдается картина митотического деления. В 4—6 нед преобладают зрелые крупные децидуальные клетки, не содержащие митозы. В них выявляются гликоген, липиды, гликозаминогликаны, кислая глицерофосфатаза, витамин С, железо, неспецифические эстеразы, дегидрогеназы янтарной и молочной кислот.
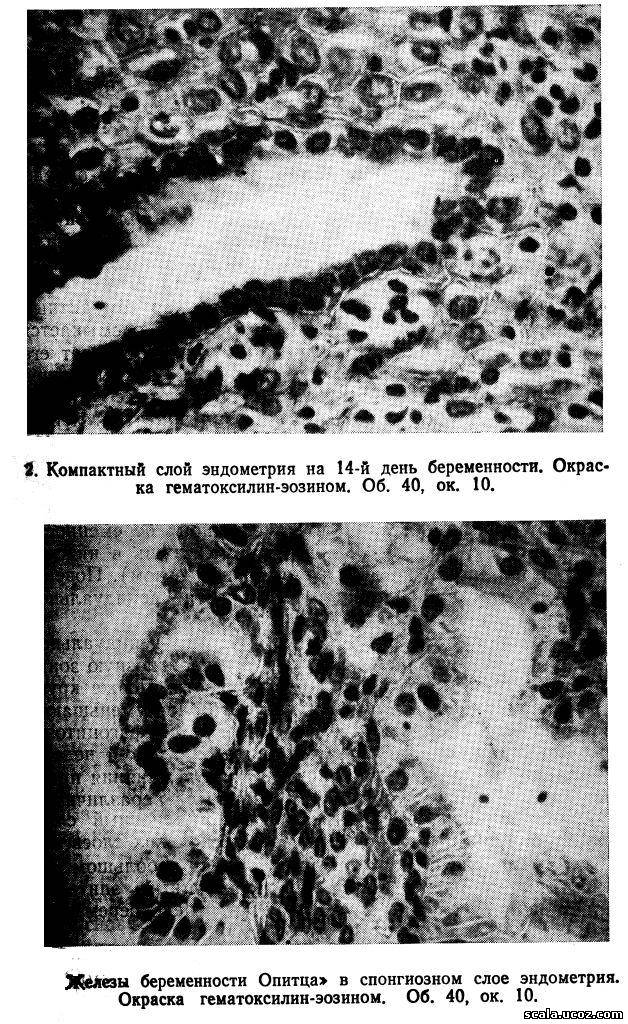
В спонгиозном слое в период имплантации железы тесно прилежат друг к другу, резко извиты, выстланы высоким эпителием, образующим сосочковые выступы, в основании которых имеется соединительная ткань. Это так называемые железы беременности Опитца (рис. 3) От желез эндометрия лютеиновой фазы менструального цикла они отличаются наличием капилляров в сосочкових выростах, окруженных аргирофильными волоконцами. Соединительная ткань, расположенная в виде тонких прослоек между железами спонгиозного слоя, не подвергается децидуальному превращению, за исключением зоны спиральных артерий.
Полная децидуализация наступает на III месяце беременности, когда толщина децидуальной ткани достигает 7 мм. У человека децидуальному превращению эндометрий подвергается на всем протяжении, но в области имплантации этот процесс происходит быстрее. В остальных отделах децидуальная ткань отчетливо определяется через 5—6 дней после завершения имплантации.
В децидуальной ткани, окружающей бластоцисту, различают капсулярную и базальную части, на остальном протяжении полость матки выстлана пристеночной децидуальной оболочкой.
Капсулярная часть, представленная компактным слоем, к 18-му дню беременности полностью смыкается над имплантированным плодным яйцом и отделяет его от полости матки. В области «пробки», закрывающей отверстие на месте имплантации, образуется «рубец», включающий клетки трофобласта, эритроциты и лейкоциты. По мере развития плодного пузыря капсулярная часть выпячивается в полость матки, закрывает ее просвет и на 16-й неделе полностью срастается с пристеночной частью децидуальной оболочки, а в дальнейшем постепенно атрофируется. При доношенной беременности капсулярная часть хорошо различима лишь в нижнем полюсе плодного яйца (над внутренним зевом]. Поверхностный эпителий в капсулярной части децидуальной оболочки отсутствует.
Утолщение пристеночной части децидуальной оболочки, включающей компактную и спонгиозную зоны, происходит до 14—15-й нед беременности. Позже крупные децидуальные клетки компактного слоя уменьшаются, частично разрушаются и подвергаются фагоцитозу. Железы спонгиозного слоя развиваются до 8-й недели беременности, затем уплощаются, а после слияния пристеночной и капсулярной части становятся неразличимы. В конце доношенной беременности пристеночный слой децидуальной оболочки состоит из нескольких слоев децидуальных клеток, содержащих в очень небольшом количестве гликоген и липиды. Поверхностный эпителий пристеночного слоя исчезает на 12-й неделе беременности.
Базальная часть децидуальной оболочки отделяет плодное яйцо от миометрия. Она состоит из компактного и спонгиозного слоев. Железы спонгиозного слоя базальной части сохраняются в большом количестве до VI месяца беременности. Постепенно они уплощаются, и в конце беременности их просвет не всегда различим. Между железами встречаются гигантские симпласты и элементы цитотрофобласта. Просвет сосудов нередко тромбирован.
В процессе имплантации продолжаются изменения в архитектонике и строении спиральных артерий, начавшиеся в предимплантационном периоде. На 7-й день беременности при имплантации зародыша в стадии бластоцисты они постепенно выпрямляются и некоторые артерии достигают поверхности эндометрия. Образуются над- сосудистые эпителиальные островки. Имплантация плодного яйца происходит в зоне этих островков с находящимися под ними спиралевидными артериями. В период нидации параллельно с децидуализацией слизистой оболочки матки артерии начинают древовидно разрастаться и достигают плодного яйца.
В периоде имплантации при погружении бластоциста в слизистую оболочку матки трофобласт продолжает пролиферировать и превращается в мощный слой тpoфобластический панцирь, состоящий из клеток кубической формы (цитотрофобласт) и расположено кнаружи синцитиальных многоядерных элементов неправильной формы (рис. 4).
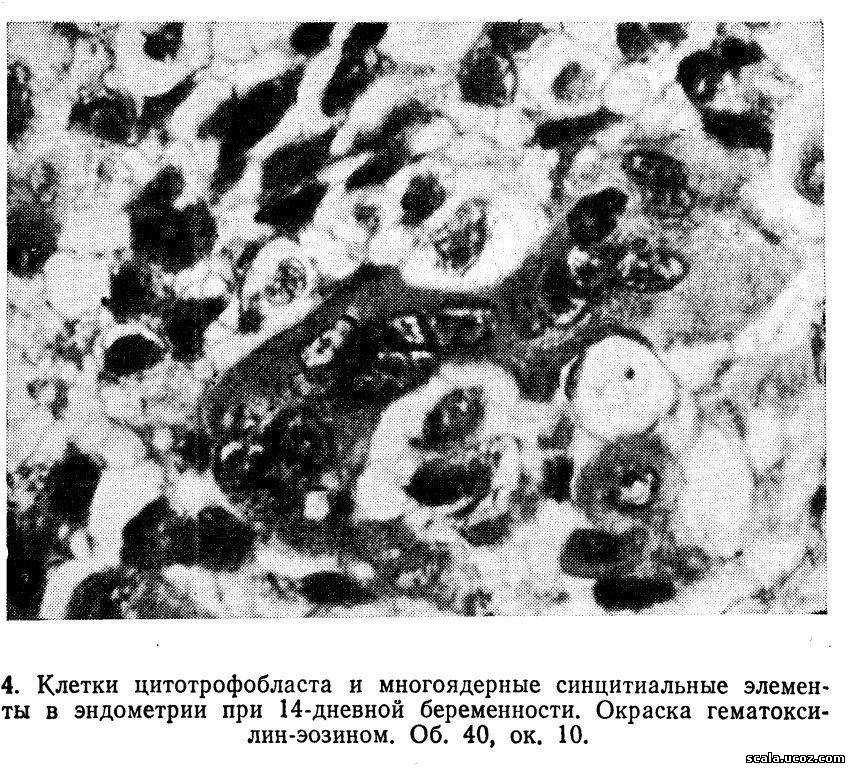
На ранних стадиях имплантадии трофобласт, не обладая выраженными цитолитическими свойствами, проникает между клетками поверхностного эпителия эндометрия, но не разрушает его. Гистолитические свойства трофобласт приобретает в процессе контакта со слизистой матки. Разрушению слизистой оболочки способствуют процессы аутолиза, связанные с активной деятельностью лизосом самого маточного эпителия. На 9-й день в трофобластическом панцыре появляются мелкие полости (лакуны), куда в результате эрозии мелких сосудов и капилляров поступает кровь матери. Тяжи и перегородки трофобласта, раздедярщие лакуны, называются первичными ворсинками, с появлением которых бластоцисту называют плодным пузырем, дальнейшем в связи со вскрытием материнский сосудов и образованием межворсинчатого пространства возникает маточно-плацентарное кровообращение. Основу первичных ворсин составляют тяжи из клеток цитотрофобласта многоугольной или неправильной формы, со светлой цитоплазмой, имеющей нежно-зернистое строение. Хорошо различимы фигуры клеточного
деления. Границы клеток, непосредственно соприкасающихся с материнской кровью, не определяются. Цитоплазма их темная, ядра удлиненной формы. Эти элементы являются предшественниками синцития неклеточного трофобласта.
В период имплантации эмбриобласт расщепляется на два листка — эктодерму, прилежащую к трофобласту, и эндодерму, обращенную в полость бластоцисты. На 8-й день из эктодермы начинается образование амниотического пузырька, дно которого является эктодермой зародыша. Сам зародыш приобретает форму щитка. На 11-е сутки в полость бластоцисты из зародышевого щитка перемещаются отростчатые клетки внезародышевой мезодермы (мезенхима). Они заполняют полость бластоцисты, окружают амниотический пузырек и выстилают трофобласт. Мезенхима, примыкающая к трофобласту, образует вместе с ним хорион. Из мезенхимы, заполняющей полость бластоцисты, и эктодермы формируется желточный мешок, дно которого, прилежащее к амниотическому пузырьку, составляет эктодерму зародыша.
Плотный тяж из клеток внезародышевой мезенхимы прикрепляет амниотический и желточный пузырьки к хориону. Это так называемая амниотическая ножка. На 15-е сутки в амниотическую ножку врастает аллантоис — пальцевидное выпячивание эктодермы заднего отдела кишечной трубки зародыша, в котором располагаются сосуды зародыша.
К концу 2-й недели беременности (12—13-й день) со стороны хориона в первичные ворсины врастает соединительная ткань — образуются вторичные ворсины. Основу их составляет соединительная ткань, а наружный покров образован эпителием (трофобласт). Они распределяются по всей поверхности плодного яйца равномерно, за исключением «фибринозной пробки», но наибольшую длину имеют в области базальной части децидуальной оболочки. Строма вторичных ворсин рыхлая, содержит аморфное вещество, в котором преобладают полисахариды и гликопротеиды, волокнистые и клеточные элементы.
В соответствии с особенностями строения стромы различают два вида вторичных ворсин [Цирельников Н. И., 1980]. Первый из них характеризуется отсутствием коллагеновых волокон, наличием тонкой сети ретикулиновых волокон и многоотростчатых клеток неправильной формы. Второй вид отличается наличием коллагеновых и ретикулиновых волокон, фибробластов и крупных клеток с большим овальным светлым ядром.
Характеризуя строму вторичных ворсин, М. Я. Субботин и В. В. Виноградов (1960) считают, что она лишь напоминает ретикулярную ткань. В действительности строма вторичных ворсин занимает промежуточное положение между внеэмбриобластической мезенхимой и синцитиализированной соединительной тканью, в которой формирование фибрилл не только связано с фибробластами, но может происходить и вне клеток [Брусиловский А. И., 1956].
Эпителий вторичных ворсин представлен двумя слоями— слоем клеток Лангханса и синцитием. Слой Лангханса состоит из клеток округлой формы со светлой цитоплазмой. Ядра их крупные, нередко наблюдаются митозы. В синцитии границы клеток неразличимы, цитоплазма темная, зернистая, с щеточной каймой. Ядра некрупные, шаровидной, овальной или цилиндрической формы.
С 3-й недели развития зародыша начинается период плацентации, который характеризуется васкуляризацией ворсин и превращением вторичных ворсин в третичные, содержащие сосуды. Процессу васкуляризации ворсин сопутствует снижение темпов роста и дифференцировки эпителия хориона.
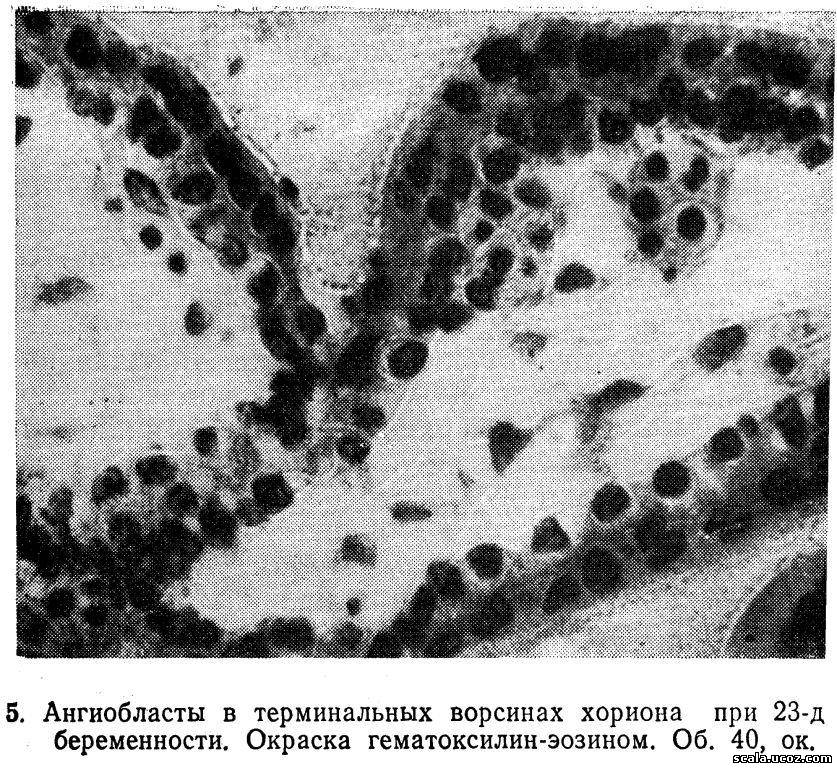
Формирование плодовых сосудов плац» происходит в процессе дифференциации ангиобластов уже у 10-дневного зародыша (Говорка Э., 1970). На 13—15-й день после оплодотворения основным группам первичных ворсин хориона сопутствуют солидные ангиобластов, в формировании которых принимают участие цитотрофобласт и мезодерма, а позже клетки коллон трофобласта. Образуются сосудистые зачатки, которые обладают способностью к самостоятельному росту вскоре превращаются в настоящие сосуды с просветом. Одновременный рост и созревание ангиобластических структур способствуют образованию сети, васкуляризирующей ворсины хориона.
Ворсины, обращенные к базальной части децидуальной оболочки, кровоснабжаются не только из с развивающихся из ангиобластов (рис. 5), но так пупочных сосудов зародыша, растущих из аллантоиса. При соединении ветвей пупочных сосудов с местной сетью кровообращения устанавливается циркуляция эмбриональной крови в третичных ворсинах, что совпадает с началом сердечных сокращений зародыша (на 21-й день развития). Формирование плодово-плацентарного кровообращения является вторым важным этапом морфогенеза плаценты.
В капсулярной части отпадающей оболочки строма гладкого хориона не содержит развитых сосудов; здесь имеются лишь их зачатки. В ветвистом хорионе капилляры, образовавшиеся во многих ворсинах, обычно располагаются непосредственно под эпителием, образуя поверхностную капиллярную сеть. Эти капилляры заполнены молодыми форменными элементами крови — мегалобластами.
Превращение вторичных ворсин в третичные является важнейшим критическим периодом в развитии эмбриона. Сущность его заключается в формировании сосудов, которое заканчивается до б—8-й, а иногда до 12— 14-й недели внутриутробного развития. По нашим наблюдениям, васкуляризация ворсин хориона заканчивается в 8—10 нед беременности, хотя отдельные бессосудистые ворсины можно встретить и в более поздние сроки. К этому времени формируется стенка сосудов, состоящая из эндотелия, базальной мембраны, мышечных волокон и соединительнотканной оболочки. Стенка капилляров представлена эндотелиальными клетками, базальной мембраной и прилежащими к ней перицитами.
В строме третичных ворсин в ранние сроки беременности отмечается высокое содержание кислых гликозаминогликанов. Степень окраски аморфного вещества наиболее высокая в центре, вокруг сосудов, а также по периферии — под хориальным эпителием.
С 8-й недели беременности в центральной части ворсин хориона выявляются пучки тонких коллагеновых волокон. На III—IV месяце содержание кислых гликозаминогликанов (гиалуроновой кислоты и хондроитинсульфата С) постепенно уменьшается и одновременно нарастает количество нейтральных гликопротеидов, белков и фибриллярных структур. Содержание хондроитинсульфата В увеличивается до 9—10-й недели беременности. Кислые гликозаминогликаны участвуют в кристаллизации фибрилл коллагена, соединении их в волокна. Формирование коллагеновых волокон связано не только с содержанием кислых гликозаминогликанов, но и с повышением активности гиалуронидазы в хориальном эпите, Последняя расщепляет гиалуроновую и хондроитинную кислоты, благодаря чему происходит их деполимеризация и уменьшается вязкость основного вещества, в свою очередь сопровождается увеличением проницаемости плацентарной ткани.
В синтезе коллагена принимает участие витамин С, большое количество которого выявлено в строме ворсин в первые 3 мес беременности. При недостаточности витамина С нарушается синтез коллагена и гликозаминогликанов. В обмене последних принимает участие витамин А.
Клеточные элементы стромы третичных ворсин представлены фибробластами и гистиоцитами. Фибробласты — отростчатые клетки веретенообразной форм слабо базофильной цитоплазмой и овальным ядром. Цитоплазма их заполнена гранулярным эндоплазматическим ретикулумом, в расширенной сети которого находятся электронно-плотное тонковолокнистое вещество, богатое белковым компонентом. Здесь много митохондрий хорошо развит пластинчатый комплекс (аппарат Гольджи), имеется большое количество митозов. Свободные клеточные элементы типа гистиоцитов крупнее, чем фибробласты, цитоплазма их интенсивно окрашена. В ранние сроки беременности в цитоплазме фибробластов и гистиоцитов содержится большое количество гликогена, кислые белки, выявлены сульфгидрильные группы.
Среди клеточных элементов стромы выделяют макрофаги (клетки Кащенко—Гофбауэра). В этих клетка является высокая активность гидролитических ферментов. В ранние сроки беременности в них найдено большое количество гликогена. Широко распространено представление о фагоцитарных свойствах этих элементов, их способности к депонированию и транспортировке тельных веществ. Для отличия некоторых форм к Кащенко—Гофбауэра от сходных с ними плазматических клеток рекомендуется окраска метиловым зеленым пиронином. При этом клетки Кащенко—Гофбауэра окрашиваются слабо, а цитоплазма плазматических клеток интенсивно окрашивается в красный цвет.
Помимо перечисленных выше клеточных элем в строме ворсин в ранние сроки беременности т опорные, или поддерживающие клетки, сходные с плазматическими, и недифференцированные мезенхимные клетки, являющиеся, по-видимому, источником образования всех клеточных форм соединительной ткани ворсин хориона.
Непосредственно к строме ворсин с наружной стороны примыкает базальная мембрана трофобласта — гомогенная эозинофильная пластинка, которая хорошо выявляется серебрением. Она состоит из трех слоев различной электронной плотности, содержит осмиофильные включения, вакуоли и соединения железа.
Хориальный эпителий васкуляризованных ворсин, как и вторичных, представлен слоем Лангханса и синцитием (рис. 6).
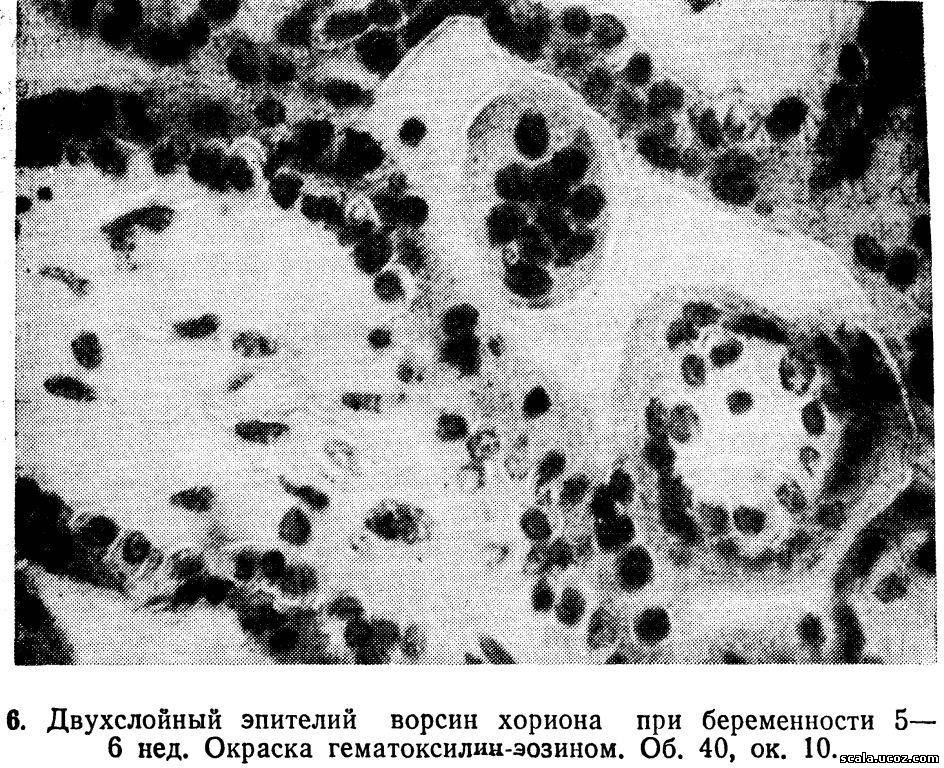
Слой Лангханса располагается на базальной мембране. Клетки цитотрофобласта имеют овальную или многоугольную форму с оксифильной цитоплазмой, они характеризуются различными вариантами распределения хроматина [Брусиловский А.И., 1976], чаще находятся в стадиях профазы и метафазы. Митотическая активность цитотрофобласта ворсин в вечернее и ночное время в 2—3 раза выше, чем в дневное, что указывает на существование определенного суточного ритма. Толщина слоя Лангханса значительно варьирует в разных ворсинах. Коэффициент вариации высоты клеток колеблется от 7,3 до 62,9 %.
Характеристика ядер цитотрофобласта в отношении содержания ДНК дана Н. И. Цирельниковым (1980). Установлено, что в 5—6 нед беременности 30—45 % ядер содержат 2 с ДНК, 10—15 % находятся в синтетической фазе, остальные ядра содержат тетраплоидное количество ДНК. В 8—9 нед уменьшается количество диплом ной и увеличивается количество тетраплоидной популяции, нарастает число ядер с межклассовым значение ДНК.
Высший пик синтеза ДНК наблюдается в конце триместра беременности, когда наиболее велико количество ядер, находящихся в синтетической фазе. В 14- 16 нед заметно падает количество клеток со значение 4 с. В целом преобладают диплоидные и тетраплоидные популяции клеток Лангханса.
В первые 4—6 нед беременности в цитоплазме клеток Лангханса много гликогена, который позже не выявляется. Липиды отсутствуют. Из ферментов в этот период найдены сукцинатдегидрогеназа (СДГ), лактатдегидррогеназа (ЛДГ), НАД-и НАДФ-диафоразы. Наиболее подробно изучена активность ЛДГ. В 5—6 нед активное ее в клетках цитотрофобласта невелика, в 8—9 нед число клеток с высокой активностью ЛДГ значительно во растает и в 11—12 нед достигает максимума. При этом происходит сдвиг активности фермента от ЛДГ-4 к Н формам.
В ранние сроки беременности цитотрофобласт третичных ворсин имеет умеренно развитую эндоплазматическую сеть, развитый пластинчатый комплекс, различное количество митохондрий, свободные рибосомы и пол сомы. Позже 8 нед количество полисом уменьшаете наблюдается увеличение пластинчатого комплекса гранулярной эндоплазматической сети. На базальной поверхности отмечаются многочисленные складки и выпячивания, в области контакта с синцитием — формирующиеся и дифференцированные десмосомы.
Кнаружи от слоя Лангханса располагается синцит альный (плазмодиальный) трофобласт—синцитий в в де непрерывного слоя базофильной цитоплазмы с включенными в нем ядрами. В зависимости от толщины сицития ядра располагаются длинной осью параллель: его свободной поверхности.
В 5—6 нед беременности синцитиотрофобласт по толщине превосходит слой Лангханса, занимая около 2/3 эпителиального пласта. Начиная с 9—10 нед синцитий постепенно истончается, а количество ядер в нем повышается.
Измерения величины плазменно-ядерных соотношениях (ПЯС) в синцитии и цитотрофобласте позволяют выделять три периода: до 8-й недели развития происходившие снипжение ПЯС синцития и нарастание ПЯС клеток цитотрофобласта, с 8-й по 12-ю неделю — стабилизация этих показателей, а в дальнейшем — снижение величин ПЯС. В синцитии преобладают ядра с диплоидным количеством ДНК.
На свободной поверхности синцития, обращенной к межворсинчатому пространству, имеются длинные тонкие многочисленные цитоплазматические выросты («щеточная кайма»), которые значительно увеличивают абсорбционную поверхность хориального эпителия.
В цитоплазме постоянно обнаруживаются липиды, железо (энзимное и транспортное), в ранние сроки беременности — гликоген, в области «щеточной каймы» — гликозаминогликаны, в цитоплазме и ядрах — кислая фосфатаза. В 8—9 нед в апикальных отделах выявляется активность щелочной фосфатазы, которая по мере прогрессирования возрастает. В синцитии присутствуют также НАД- и НАДФ-диафоразы, 5-нуклеотидаза, СДГ и ЛДГ. В 5—6 нед беременности обнаруживаются интенсивно окрашенные гранулы формазана, что свидетельствует о высокой активности ЛДГ. 3. П. Жемкова и О. И. Топчиева (1973) отмечают общее повышение ферментативных реакций в синцитии ворсин на 2—3-м месяце развития плода. Наличие большого количества РНК и белков в эти сроки связывают с активацией синтеза белка и гонадотропинов. Начиная с 3-го месяца внутриутробного развития активность хориального эпителия постепенно снижается.
Окончательная дифференцировка хориального эпителия заканчивается на 6—7-й неделе эмбриогенеза, что по времени соответствует редукции желточного мешка и переходу с желточного на аллантоисное кровообращение.
При электронно-микроскопическом исследовании между синцитием и трофобластом найдены щелевидные («субсинциатиальные») пространства. С помощью специальных ходов они имеют сообщение с базальной мембраной трофобласта, что обеспечивает возможность контакта между синцитием и стромой ворсин в ранние сроки беременности, когда непосредственная связь между этими структурами отсутствует.
В ранние сроки онтогенеза число свободных рибосом в синцитии очень невелико. Основная часть их связана с мембранами эндоплазматической сети. Имеются редкие полисомы и мелкие лизосомы. Митохондрии мелкие, многочисленные. Постоянно выявляются секреторные и осмиофильные гранулы. Цитоплазма базальной поверхности синцития образует многочисленные тонкие выпячивания, соприкасающиеся с клетками Лангханса.
При изучении ультраструктуры третичных ворсин в ранние сроки беременности выделены зоны определенной дифференцировки синцития в направлении от апикальной к базальной поверхности: наружная — зона абсорбции, средняя — секреторная зона, базальная — зона накопления (с преимущественным нахождением гликогена и митохондрий). На основании электронно-микроскопической характеристики выделены также участки синцития с различной функциональной активностью.
Источником образования синцитиотрофобласта ворсин хориона служат клетки слоя Лангханса. С помощью электронной микроскопии различают типы клеток цитотрофобласта, отражающие стадию их дифференцировки и превращения в синцитий. Можно предположить, что не все клетки Лангханса трансформируются в синцитий. Часть таких клеток, обладающих характерной структурой, служит источником образования клеточных колонн и островков.
Клеточные колонны — это тяжи цитотрофобласта, расположенные в дистальных отделах ворсин и соединяющие соседние ворсины. Они хорошо различимы до IV месяца беременности. С помощью клеточных колонн часть ворсин, достигающих децидуальной ткани, закрепляется в компактном слое. На поверхности децидуальной базальной оболочки цитотрофобласт образует базальную пластину. В области соприкосновения трофобласта и децидуальной ткани формируется зона коагуляционного некроза (слой Нитабух), которая входит в состав базальной мембраны.