Обмен газов в плаценте аналогичен газообмену в легких и определяется прежде всего наличием плацентарного кровообращения. Доказано что проникновение кислорода (О2), углекислоты (С02), наркотических газообразных соединений происходит за счет физико-химических процессов осмоса и диффузии.
Плацента не обладает способностью к накоплению О2 и СО2, поэтому транспорт их происходит постоянно. Плацентарный барьер является фильтром, пропускающим газы в количестве, прямо пропорциональном их концентрации и обратно пропорциональном толщине мембраны. Участки ворсинчатого хориона, покрытые микроворсинами, служат местом транспорта О2 и СО2. При этом эритроциты непосредственно прилегают к микроворсинам [Ibrahim М. Е. et al., 1982]. Площадь пограничной поверхности ворсин, определяющая условия газообмена плода на 1 кг его массы, более чем в 3,5 раза превосходит площадь поверхности легочных альвеол (на 1 кг массы тела взрослого) и составляет 13 м2, или 3,4 м2/кг.
Через мембрану капилляра кислород и углекислота проникают, только будучи растворенными в плазме крови. Имеется линейная зависимость между количеством растворенного в крови кислорода и напряжением его в крови (Ро2), т. е. количество 02, растворенного в крови, при данной температуре прямо пропорционально Ро2. При растворении в 100 мл крови 0,235 мл О2 (при температуре 37 °С) Ро, составляет 100 мм рт. ст. Вдыхание 100 % кислорода может увеличить Ро2 в крови до 700 мм рт. ст., что позволяет транспортировать 1,4 мл кислорода в 100 мл крови.
Газообмен в плаценте характеризуется двумя важнейшими процессами: проникновением О2 и выведением СО2. Эти процессы можно рассматривать как два взаимопротивоположных каскада парциальных давлений этих газов.
Газообмен между организмами матери и плода осуществляется как через плаценту, так и через околоплодные воды. Трансплацентарный обмен 02 и С02 обусловлен многими факторами. Он зависит от количества кислорода, поступающего в матку, состояния маточноплацентарного и фетоплацентарного кровотока, включающего сосуды пуповины. Распределение кислорода, поступающего в матку, осуществляется достаточно закономерно.
S. L. Romney и соавт. (1955) во время операции кесарева сечения vcтлпопили, что в 100 мл крови, полученной из маточной артерии, содержится 15,2 вд кисдорода, в крови из маточной вены— 10,5 мл. Артериовенозная разница составляет 4,7 мл в 100 мл крови. Следовательно, матка в час потребляет 2100—2250 мл кислорода. Этот кислород частично утилизуется в процессе метаболизма в самом миометрии, но большая часть его поступает в плаценту и потребляется плодом и плацентой. Ткани плаценты утилизуют около половины проникающего в них кислорода [Dawes G. S., 1971; Meschia G., 1978). При беременности 3 мес плацента потребляет кислорода 252 мл/(кг-ч), в то время как к плоду он поступает в количестве 90—138 мл/(кг-ч). Зрелая плацента потребляет кислорода около 198 мл/(кг-ч), что в 2—3 раза больше потребления кислорода тканями плода. В случае кислородного голодания плода отмечаются снижение потребления кислорода плацентой и преобладание в плаценте процессов анаэробного гликолиза.
В межворсинчатом пространстве содержится 40— 48 мл кислорода. Помимо этого, существует так называемый перфузионный резерв плаценты, равный 50 % минутного объема крови, притекающей к плаценте, который представляет собой определенный запас кислорода для обеспечения потребностей плода в экстремальных условиях. Во время схватки давление в межворсинчатом пространстве возрастает вдвое, однако количество крови в нем остается неизменным и содержание кислорода существенно не меняется.
Достаточное насыщение плацентарной крови кислородом и, следовательно, адекватное снабжение им плода зависят от Ро2 и содержания гемоглобина в крови матери, а также от состояния маточно- и фетоплацентарного кровообращения. Так, при величине гемоглобина 150 г/л содержание кислорода в крови приближается к 1600 мл, при 100 г/л —к 1200 мл и при 50 г/л —всего к 900 мл кислорода [Sallivan S. F., 1972]. Кислород из крови матери переходит в кровь плода через стенки сосудов в ворсинах хориона. Газообмен в системе мать - плод нельзя свести к механизму простой диффузии кислорода и углекислоты в результате градиента (различия концентрации и давления) Ро2 или Рсо2. Безусловно, в плаценте имеет место диффузия газов, их переход осуществляется за счет разности напряжения кислорода по обе стороны плацентарного барьера, которая составляет в среднем 70 мм рт. ст. В то же время известно, что в Условиях плацентарной недостаточности кислородное голодание v плода возникает лишь при уменьшении разницы между Рош в крови матери и плода только Дб 8—10 мм рт. ст. [Seeds А. Е., 1980]. Следовательно, ве-личина градиента Ро2 не играет ведущей роли в диффузии кислорода через плацентарную мембрану.
В транспорте кислорода и углекислоты, помимо состояния структурных элементов в плацентарной мембране, имеют значение толщина ее и ряд других факторов. К ним прежде всего относятся приспособительные механизмы в организме плода: большой минутный объем сердца, высокая скорость кровотока, наличие в крови фетального гемоглобина, полицитемия и преобладание анаэробных процессов метаболизма. Для транспорта кислорода и углекислоты весьма важна также степень диссоциации кислорода в крови матери и плода.
Так, кровь плода может транспортировать О2 в количестве 23% по объему, в то время как кровь матери — только 16 % по объему. При одинаковом содержании кислорода величина Ро2 в крови плода всегда ниже, чем в крови матери. Это вызывает изменение градиента давления кислорода в сторону передачи его из материнской крови в кровь плода. Этому способствует концентрация гемоглобина в крови плода, которая почти на 50 % выше, чем в крови матери, а также увеличенное количество эритроцитов. Полицитемия ведет к снижению давления кислорода в крови в капиллярном русле ворсин хориона и способствует его транспортировке.
Существует тесная связь газообмена в плаценте с процессами метаболизма в организме плода, в частности с накоплением кислых продуктов обмена в его крови (Н+, лактат, С02). Так, передаче кислорода плоду способствует иная (смещение кривой диссоциации влево) диссоциация кислорода под влиянием ацидоза в его крови и изменения величины рН. Обычно при более высокой концентрации Н+ в крови сродство кислорода к гемоглобину уменьшается, а передача и присоединение кислорода к гемоглобину замедляется, при этом происходит освобождение кислорода. С другой стороны, уменьшение концентрации Н+ и соответственно увеличение рН крови повышают сродство кислорода к гемоглобину с образованием комплекса НС02. В плаценте этот эффект выражен особенно резко.
На рис. 18 представлена схема перфузионной системы с функциональными шунтами материнской части плаценты. По артериям пуповины в плаценту попадают кровь с высоким содержанием С02 и лактата, которые через плацентарную мембрану переходят в межворсинчатое пространство. После того как СО2 и связанные кислоты (лактат и др.) выходят из крови плода, величина рН ее возрастает и увеличивается возможность перехода кислорода в кровь плода из межворсинчатого пространства. Одновременно в крови матери повышение содержания СО2 и кислот приводит к накоплению кислых продуктов и уменьшению рН (ацидоз). Это вызывает высвобождение кислорода и создает предпосылки для его перехода через плацентарную мембрану.
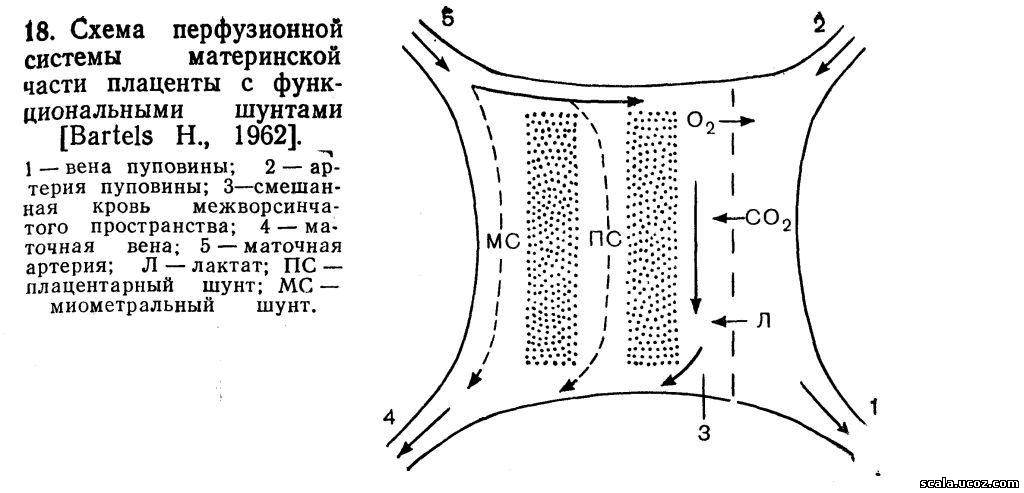
В крови, оттекающей от плаценты к плоду, из вены пуповины насыщение кислорода (нас. О2) и Ро2 значительно ниже, чем в крови не только из маточно-плацентарных сосудов, но также в капиллярной и венозной крови матери (табл. 6). Артериовенозная разница (АВР) в крови сосудов пуповины незначительно отличается от таковой в периферической крови матери, но оказывается существенно ниже, чем в крови маточных сосудов, где артериовенозная разница по Po2 составляет около 70 мм рт. ст., а по нас. 02 —около 40 %. Это свидетельствует о том, что, несмотря на почти одинаковое и даже несколько большее потребление кислорода организмом плода по сравнению с материнским, в плаценте процесс переноса кислорода протекает не только по закону простой диффузии, обусловленной величиной градиента концентрации.
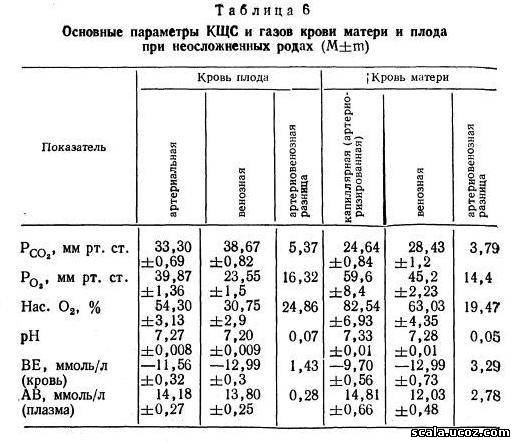
Транспорт углекислоты через плаценту имеет особенности. Величина Рсо2 в венозной и артериальной крови пуповины обычно бывает выше, чем в периферической крови матери. Плодово-материнский градиент (ДРсо2 ) в артериальном и венозном бассейнах крови является положительным и составляет +8,66 и + 10,24 мм рт. ст. Однако у некоторых рожениц наблюдаются отрицательные значения ДРсо,» что указывает на возможность транспорта СО2 не только в направлении от плода к матери, но также из организма матери к плоду.
Транспорт углекислоты тесно связан с переносом не только кислорода, но и бикарбонатов (НС0-з), кислых продуктов обмена, глюкозы и других метаболитов и совершается не только путем простой диффузии. Концентрация бикарбонатов (АВ) в крови матери и плода практически одинаковая, что является следствием свободного двустороннего трансплацентарного перехода этих веществ из организма матери к плоду и наоборот (мать-плод). Следует отметить, что и количество кислых продуктов обмена (BE) одинаково в венозной крови матери и плода. Это также обусловлено свободным их переходом через плацентарную мембрану. В связи с этим можно предположить, что метаболический ацидоз и положительный ДРсо2, по-видимому, являются ведущими в трансплацентарном газообмене.
Исследования газового состава и величины рН околоплодных вод показали существование параплацентарного газообмена между организмом матери и плода.
Вопрос об участии амниотической жидкости в снабжении плода кислородом возник тогда, когда впервые были получены сведения о наличии в них растворенных газов. Однако на него был дан отрицательный ответ, поскольку величины Ро2 , полученные с помощью полярографического метода, были значительно ниже таковых крови плода. Тем не менее было выявлено, что концентрация кислорода в амниотической жидкости не только коррелирует с Po2 кожных покровов плода, но и зависит от напряжения кислорода в тканях матки и плаценты.
Полярографические исследования тканевого Ро2 в системе мать—плацента—плод в условиях эксперимента У беременных кроликов позволили уточнить роль околоплодных вод в снабжении плода кислородом (табл. 7).

Как видно из таблицы, наибольшая величина Ро, определяется в подкожной жировой клетчатке беременных кроликов, на втором месте находится миометрий, затем следуют околоплодные воды и мозг плода. Наименьшие показатели Ро2 выявлены в плаценте. Наличие относительно высокого уровня Ро3 в околоплодных водах, практически не отличающегося от напряжения кислорода в миометрии, указывает на активный переход кислорода в полость амниона, по-видимому, через внеплацентарную часть плодных оболочек.
Представляют интерес результаты исследования Ро 2 в ткани плаценты. Известно, что маточно-плацентарная циркуляция в течение беременности является наиболее интенсивной частью гемодинамики матки, однако в плацентарной ткани определялось низкое напряжение кислорода, которое варьировало от 26,5 до 6,5 мм рт. ст. Градиент снижения величины Ро8 в направлении миометрий-околоплодные воды-плацента имел место в 2/3 плодовместилищах. Это свидетельствует о высокой потребности в кислороде и активности окислительных процессов в плаценте, поскольку Ро2 тканей отражает не только поступление, но и утилизацию кислорода тканями, а также указывает на меньшую вероятность перехода кислорода в околоплодные воды через амнион плаценты.
Практически одинаковые значения Ро2 околоплодных вод и тканей плода свидетельствуют о возможности участия их в снабжении плода кислородом. Параллельные клинические исследования на аппарате микро-Аструп (фирма «Biomedica», Дания) параметров КЩС и газов крови матери, плода (кровь из сосудов пуповины забирали в момент рождения ребенка до его первого вдоха) и околоплодных вод, проведенные в конце беременности и в родах, подтверждают участие околоплодных вод в газообмене плода (табл. 8).

Как видно из табл. 8, во всех наблюдениях, кроме группы рожениц с нефропатией, Ро2 околоплодных вод было значительно выше, чем в капиллярной крови матери и крови из вены пуповины. Следовательно, плод находится в среде с высоким напряжением кислорода.
При этом можно думать о нескольких путях воздействия кислорода амниотической жидкости на газообмен плода (чрескожный, через желудочно-кишечный тракт и легкие).
Механизм транспорта кислорода в околоплодные воды практически не изучен. В первую очередь нельзя исключить наличия простой диффузии из микрососудов децидуальной оболочки, поскольку величины Ро, амниотической жидкости и артериальной крови, поступающей в миометрий, близки. Нельзя также исключить участие липидов водной оболочки в активном транспорте О2 между организмом матери и плода. В литературе имеются также сведения о том, что растворимость О2 в липидах в десятки раз выше, чем в водных растворах. Это обусловливает взаимодействие клеточных мембран и цитоплазмы с кислородом.
Механизм кинетического и кинетически-диффузионного переноса кислорода в жидкостях и тканях организма изучен достаточно подробно [Березовский В. А., 1972]. Установлено, что кинетический перенос кислорода обычно опережает диффузионную его доставку. Так, молекулы воды свободно проходят через стенку капилляров, а вместе с ней проникает и растворенный в воде кислород. Транспорт кислорода на отдельных участках осуществляется через межклеточные щели, которыми очень богаты плодные оболочки, заполненные межтканевой жидкостью. Через клеточные мембраны кислород может проникать, если диаметр пор составляет 60— 70 нм.
Как показали наши исследования, ингаляции 60 % кислородно-воздушной смеси увеличивают тканевое Ро2 и содержание кислорода в крови матери на 200 % и более. Одновременно наблюдается повышение Ро2 (и 2 раза и более) и в околоплодных водах. Это становится возможным лишь при быстром транспорте кислорода в околоплодные воды через всю поверхность плодных оболочек.
Прямой зависимости величин Ро2 от рН и Рсо2 в околоплодных водах не выявлено. Однако во всех наблюдениях с высокими значениями рН околоплодных вод (более 7,2) Ро, превышает 100 мм рт. ст. У беременных и рожениц с низким Рсо2 вод (до 30 мм рт. ст.) напряжение кислорода высокое (115,42±5,43 мм рт. ст.], что, по-видимому, связано с меньшим выведением углекислоты из организма плода в результате меньшей продукции ее в условиях высокой концентрации кислорода в около-плодных водах. Подтверждением этого служат результаты сравнительного изучения Ро2 околоплодных вод Н Рсо2 крови пуповины: у плодов при Ро2 в околоплодных водах свыше 100 мм рт. ст. в крови пуповины отмечается самая низкая концентрация углекислоты (23,62± ±2,9 мм рт. ст.). В то же время при Ро2 в околоплодных водах от 80 до 100 мм рт. ст. Рсо2 в пуповинной: крови составляет 31,33+1,04 мм рт. ст. (р<0,05), а при.. Ро2 до 80 мм рт. ст. Рсо2 в пуповинной крови равно.] 34,62±3,64 мм рт. ст. (р<0,05). Следовательно, напряжение кислорода в околоплодных водах влияет на продукцию углокислоты в организме плода и на концентрацию ее в пуповинной крови.
Помимо этого, в серии других экспериментальных исследований нами получены несомненные доказательства снабжения плода кислородом через околоплодные воды [Савельева Г. М. и др., 1984]. Исследование было проведено на плодах кроликов, находящихся в условиях с различным напряжением кислорода в околоплодных водах при полном прекращении маточно-плацентарного кровообращения. Нарушение маточно-плацентарного кровообращения и острую тяжелую гипоксию у плода создавали путем извлечения неповрежденного плодного яйца (включающего плаценту, интактные оболочки, воды и плод) из рога беременной матки кролика и погружения его на 9 мин в ванночку с водой при температуре 38—39°С. Операцию производили у крольчих под нейролептаналгезией (натрия оксибутират по 0,5 мг/кг и дроперидол по 0,5 мг/кг) в сочетании с местной инфильтрационной анестезией 0,25 % раствором новокаина.
Различного напряжения кислорода в околоплодных; водах достигали введением в полость амниона разных плодов по 3 мл «нормоксического» (со стандартным Ро2 —158 мм рт. ст.), «гипероксического» изотонического раствора натрия хлорида (при Ро2 300 мм рт. ст.) или аноксической смеси (при нулевом значении Ро2 ) изотирчікого раствора натрия хлорида с натрия сульфатом. Растворы готовили непосредственно перед операцией, контроль Ро2 в них проводили на полярографе.
Состояние плода оценивали по величинам рН и Ро2 крови, полученной по первого его вдоха, предварительно определив те же параметры в околоплодных водах, после забора их в стеклянные капилляры в анаэробных условиях. Эти исследования проводили на аппарате микро-Аструп.
Для проведения сравнения результатов исследования в каждую серию было включено по 5 плодов из одного рога беременной матки крольчихи, которые извлекали в неповрежденном плодном яйце последовательно с интервалом в 1 мин. В амниотическую полость трех первых плодов сразу после отделения от организма матери и перед погружением в ванночку вводили одну из приготовленных жидкостей: аноксическую, нормоксическую или гипероксическую. Четвертое плодное яйцо погружали в ванночку без предварительного введения изотонического раствора натрия хлорида. Спустя 9 мин проводили забор околоплодных вод и крови плодов для соответствующего анализа. Контролем служил здоровый пятый плод; околоплодные воды и кровь получали сразу же после извлечения его из матки.
В результате проведенных экспериментов было установлено следующее. У здоровых плодов Роа околоплодных вод (79,1+3,4 мм рт. ст.) в 2—3 раза превышало Ро, их крови (28,7+2,48 мм рт. ст.). Такое же соотношение напряжения кислорода в околоплодных водах и в крови плода наблюдалось во всех исследуемых группах, причем в околоплодных водах у здоровых плодов Ро2 варьировало от 65 до 102 мм рт. ст. Следовательно, высокий уровень кислорода в околоплодных водах был обусловлен переходом его в полость амниона из крови матери. В артериальной крови (из маточной артерии) беременных крольчих во время операции Р02 находилось на высоком уровне и варьировало от 142 до 102 мм рт. ст., составляя в среднем 122+6,8 мм рт. ст. при величине рН 7,34+0,02.
При тяжелой гипоксии плода, вызванной отделением плодного яйца от материнского организма, отмечается Умеренное окрашивание околоплодных вод меконием, Угнетение двигательной активности и рефлекторной деятельности плода. Напряжение кислорода в крови плодов при гипоксии существенно (р<0,001) отличается от такового здоровых плодов, так же как рН их крови (Р < 0,001). В околоплодных водах Ро2 в среднем становится на 15,4% ниже (р<0,05), чем в группе здоровых плодов, что, по-видимому, связано с утилизацией кислорода из амниотической жидкости. Значительно снижается величина рН околоплодных вод, так же как в крови плода при гипоксии, за счет выведения в амниотическую жидкость из организма плода большого количества кислых продуктов обмена (табл. 9).
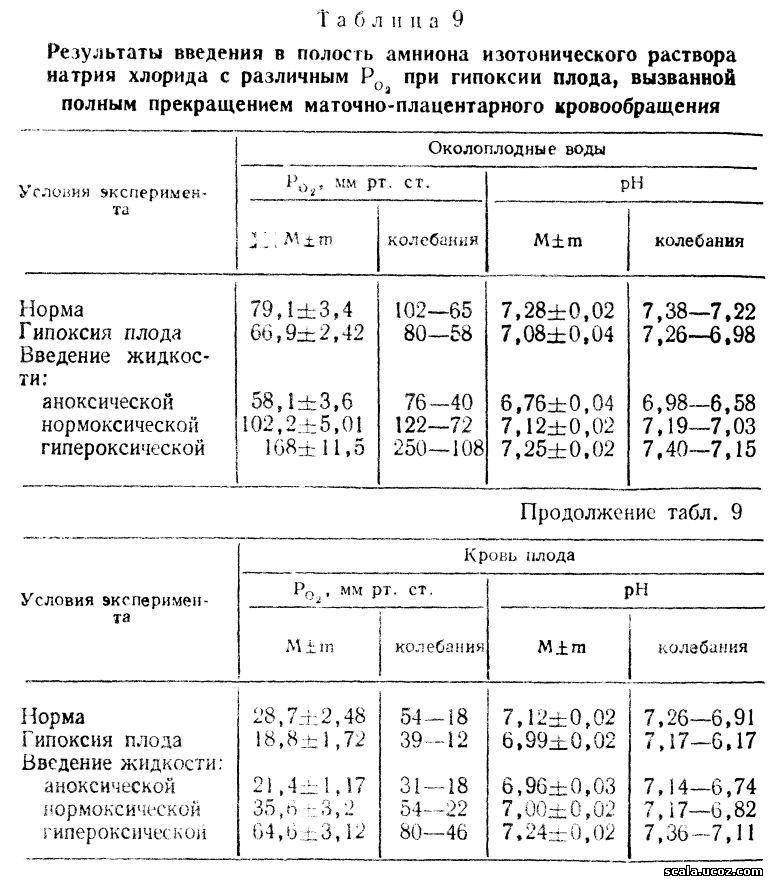
Введение аноксической смеси в полость амниона вызывает уменьшение Р02 в околоплодных водах, однако на объективных показателях оценки состояния плода это практически не отражается. Клинически выявляется более значительное окрашивание меконием околоплодных вод. Практически отсутствует двигательная активность плода. О тяжести состояния плода свидетельствует величина рН околоплодных вод, составляющая 6,76± ±0,04, которая чрезвычайно низка и существенно (р <0,001) отличается от рН вод в предшествующих группах плодов.
Следовательно, нарушение матичиоилацентарного кровообращения и прекращение трансплацентарного снабжения плода кислородом приводит к развитию тяжелой гипоксии плода, о чем свидетельствует выраженный ацидоз в крови и гипоксемия плодов, при этом в околоплодные воды выводится значительное количество кислых продуктов обмена. Выживаемость плода в течение 9—10 мин обусловлена высоким напряжением кислорода в околоплодных водах. Индивидуальный анализ наблюдений показал однонаправленность изменений изучаемых параметров: чем меньше напряжение кислорода в амниотической жидкости, тем ниже величина рН, что указывает на выраженное страдание плода.
Интересные данные получены в наблюдениях с увеличенным и чрезмерным содержанием кислорода в полости амниона изолированного плодного яйца. Введение в полость амниона нормоксической жидкости (Ро. 158 мм рт. ст.) существенно повышает содержание кислорода в околоплодных водах. Так, спустя 9 мин после изоляции плодного яйца Ров в околоплодных водах стало в 1,5 раза выше, чем у плодов, оставшихся без лечения, в состоянии гипоксии, и в 1,3 раза выше, чем в водах здоровых плодов (р<0,001). Существенно выше оказалась и величина рН околоплодных вод (7,12± ±0,02) по сравнению с двумя предшествующими группами, несмотря на то, что рН крови плодов (7,0±0,02) остается таким же, как и плодов без оксигенотерапии. Обращает на себя внимание, что напряжение кислорода (35,6±3,2 мм рт. ст.) в крови плодов было существенно выше Ро2 крови плодов предшествующих групп, в том числе и здоровых плодов. По-видимому, это про-исходило за счет утилизации кислорода из околоплодных вод. Об этом же свидетельствует и относительно большая величина рН амниотической жидкости в результате меньшей экскреции плодом кислых продуктов обмена.
Массивная оксигенация околоплодных вод существенно изменяет величины исследуемых параметров. Так, через 9 мин после введения гиперокоической смеси (при Ро, около 300 мм рт. ст.) в полость амниона Ро2 околоплодных вод становится в 2,12 раза выше нормы и составляет 1684 ± 11,5 мм рт. ст., величина рН (7,25 ± 0,02) соответствует норме, а в некоторых наблюдениях достигает даже 7,4. Особенно разительные результаты получены при исследовании крови плода. Имеет место очень высокое, нехарактерное для экспериментальных животных напряжение кислорода (64,64 ± 3,12 мм рт. ст.) в крови плодов, которое в среднем в 2,4 раза выше нормы. Величины рН крови (7,24±0,02) также значительно превышают таковые здоровых плодов. Высокие показатели рН крови плодов и амниотической жидкости свидетельствуют о высоком уровне обменных процессов, что способствует выживанию плода в условиях нарушенного маточно-плацентарного кровообращения. Клинически у этих плодов через 9 мин после прекращения плацентарного кровотока сохранялась хорошая двигательная активность и рефлекторная деятельность.
Нами установлена одинаковая направленность изменений Р02 и рН крови плода в зависимости от напряжения кислорода в околоплодных водах (рис. 19). Наиболее выраженный ацидоз в крови плода наблюдается при введения аноксической жидкости в полость амниона. Под влиянием повышенного содержания кислорода околоплодных вод, даже несмотря на длительную изоляцию плодного яйца от материнского организма, параметры жизнедеятельности и метаболизма плода указывают не только на отсутствие его страдания, но и на высокую океигенацию его организма. Устанавливается равновесие в обмене кислыми и щелочными продуктами между плодом и амниотической жидкостью.
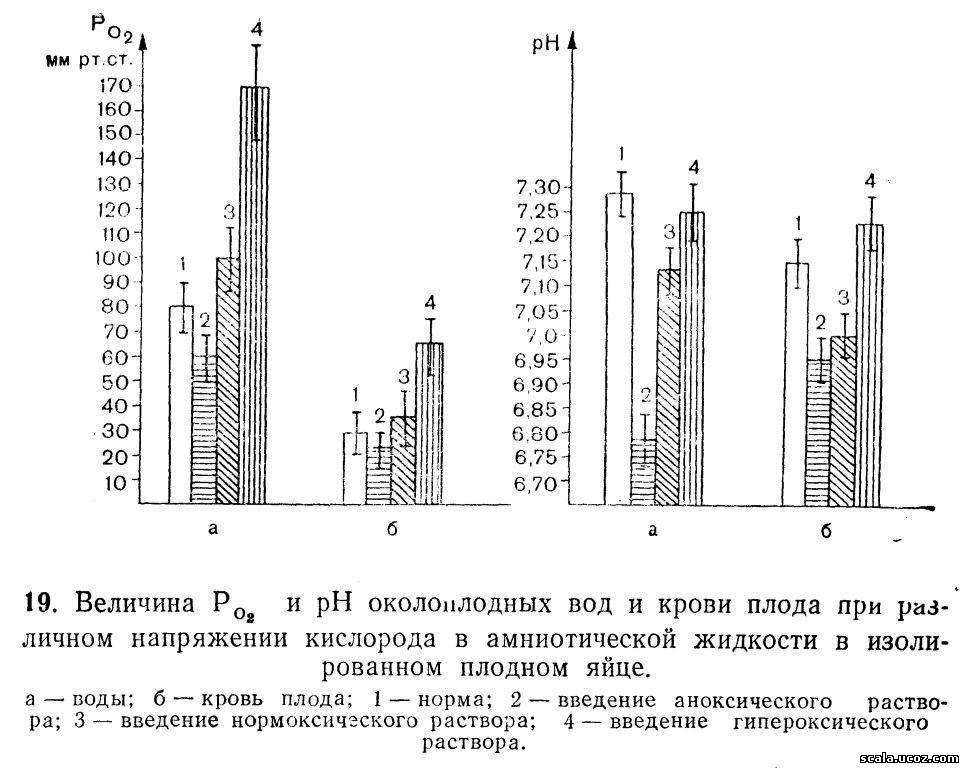
Таким образом, полученные результаты свидетельствуют о снабжении плода кислородом не только плацентарным путем, но и через околоплодные воды.
Углекислота выводится в организм матери от плода не только через сосуды пуповины в плаценту, но и в околоплодные воды. При этом накопление ее в организме плода практически не происходит. В околоплодных водах величина Рсо2 существенно выше, чем в крови матери и плода, и градиент снижения концентрации углекислоты имеет направление воды -плод-мать (табл. 10).
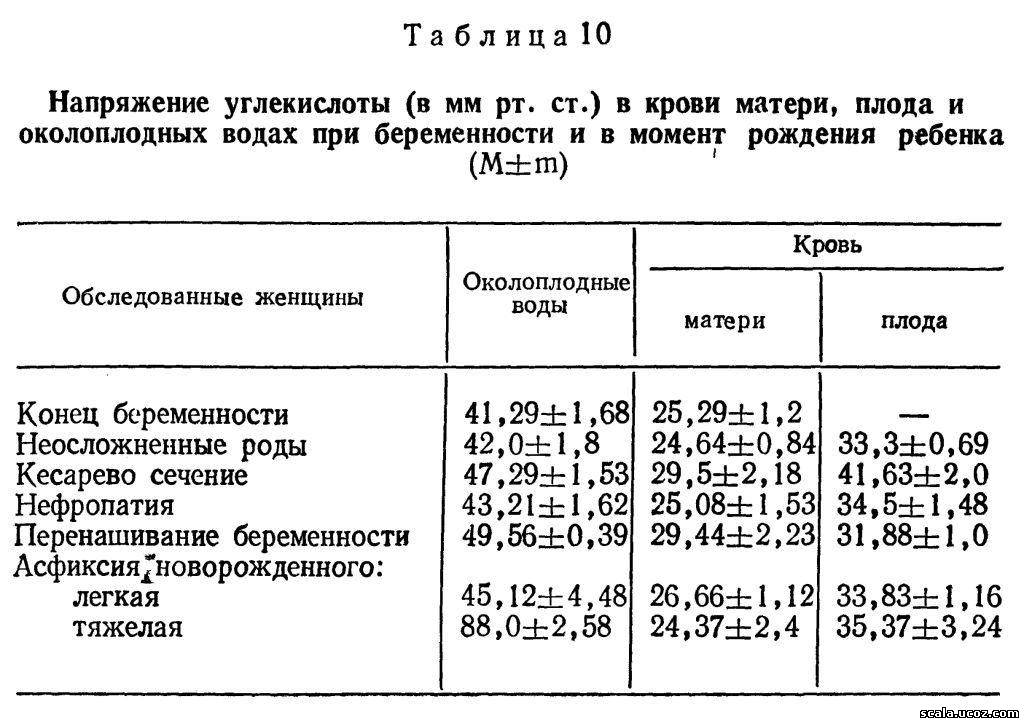
Как видно из таблицы, наиболее низки величины Рсо2 в околоплодных водах в конце не осложненной беременности. Существенно выше показатели Рсо2 при перенашивании беременности и особенно при тяжелой гипоксии плода. Накопление углекислоты в околоплодных водах не сопровождается изменением ее концентрации ни в крови плода, ни в крови матери, а вызвано преобладанием трансамниального ее выведения из организма плода, по-видимому, в связи с затрудненным прохождением ее через пуповину в плаценту. Характерно, что трансамниальный газообмен протекает чрезвычайно быстро (2—3 мин) в связи с высокой липотропностью кислорода и углекислоты [Seeds А. Е., 1980].
Следовательно, между организмом матери и плода существует сложная динамичная система газообмена, механизмы которой еще полностью не изучены.