Морфологические компоненты коменсаторно-приспособительных реакций плаценты обычно описывают в виде гиперплазии резорбционных ворсин, синцитиотрофобласта и капилляров хориона, развивающейся в условиях гипоксии при разнообразных патологических воздействиях [Пузанкина Е. Б., 1972; Жемкова 3. П. и Топчиева О. И., 1973, и др.].
Исходя из основных законов биологии развитие фетоплацентарного комплекса, как и любой живой системы, нельзя рассматривать вне приспособительных реакций, позволяющих сохранять целостность организма при изменении условий внешней среды в норме и патологии. Следовательно, формирование компенсаторно-приспособительных механизмов является физиологическим процессом.
Исходя из основных законов биологии развитие фетоплацентарного комплекса, как и любой живой системы, нельзя рассматривать вне приспособительных реакций, позволяющих сохранять целостность организма при изменении условий внешней среды в норме и патологии. Следовательно, формирование компенсаторно-приспособительных механизмов является физиологическим процессом.
В связи с тем что основные процессы, поддерживающие гомеостаз системы мать—плацента—плод, происходят в структурах, разделяющих кровь матери и плода, морфологические проявления компенсаторно-приспособительных реакций имеют значение прежде всего в области плацентарного барьера.
Плацентарный барьер состоит из тканей плодного происхождения (трофобласт, базальная мембрана трофобласта, соединительнотканная строма ворсин, базальная мембрана эндотелия капилляров и эндотелиальные клетки) .
Ворсины малого калибра и терминальные являются областью обмена между кровью матери и плода. Крупные ворсины служат скелетом, несущим опорную функцию и функцию регуляции поступления фетальной крови в капилляры мелких ворсин.
Биометрический анализ компонентов плацентарного барьера в I триместре беременности, проведенный А. И. Брусиловским, показал, что толщина эпителиального покрова ворсин составляет 30 усл. ед. (коэффициент вариации 23,5 %). Толщина синцития в среднем равна 14,2±0,80 усл. ед., цитотрофобласта — 17,5±0,80 усл. ед. Цитотрофобласт толще синцития в среднем на 7,2 усл. ед.
Средняя толщина стенки функционирующих сосудов составляет 23,0±1,43 усл. ед. В I триместре беременности еще мало капилляров, расположенных в периферической части ворсин. Больше всего сосудов (48 %) находится в средней зоне ворсины, на расстоянии от 30 до 80 усл. ед. от мембраны трофобласта. В центральной зоне, на расстоянии более 80 усл. ед. от мембраны трофобласта, располагается 37% сосудов. Лишь 15% сосудов обнаруживается в подцитотрофобластической зоне (на расстоянии 20,5 усл. ед. от трофобласта).
Во второй половине беременности происходит истончение плацентарного барьера, которое дает возможность максимального сближения русла кровообращения матери и плода. Этот процесс обусловлен исчезновением клеток цитотрофобласта, которые служат источником образования синцития, уменьшением количества клеточных элементов стромы ворсин и перемещением капилляров к периферии ворсин (в подцитотрофобластическую зону).
При доношенной беременности плацентарный барьер состоит из 7 электронно-микроскопических слоев (рис. 15): синцитиотрофобласта, светлого пространства, двух базальных мембран трофобласта и коллагеновой ткани между ними, базальной мембраны эндотелия капилляра и эндотелия плодового капилляра [Snoeck J., 1956].
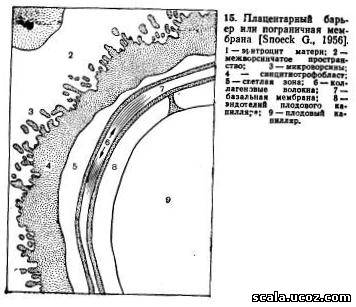
Ядра в синцитиотрофобласте располагаются неравномерно, часть из них — группами. Имеются участки, лишенные ядер, примыкающие непосредственно к стенке капилляра. В этих областях наиболее интенсивно происходит обмен между организмами плода и матери. В синцитии, помимо микроворсинок, найдены цитоплазматические выросты, обращенные к базальной мембране, которые считаются зонами активного всасывания.
При исследовании ультраструктуры эндотелия капилляров выявлено наличие в них микрофиламентов, располагающихся в большом количестве в цитоплазме. Их рассматривают как контрактильные элементы и обнаруживают чаще при доношенной беременности.
В зрелой плаценте в 60 % ворсин толщина синцития меньше, чем в период плацентации, и составляет 12,5± ±0,90 усл. ед. Значительно тоньше становится стенка сосудов (7,3±0,58 усл. ед.). Максимальное расстояние сосудов от базальной мембраны составляет 60 усл. ед. На расстоянии 3—9 усл. ед. находится до 56 %; а на расстоянии до 3 усл. ед. — 20 % сосудов. Значительная толщина барьера в незрелой плаценты. Удаление сосудов от базальной мембраны указывают на меньшую возможность осуществления процессов плодово-материнского обмена по сравнению со зрелой плацентой. В связи с этим в незрелой плаценте особенно велика потребность в активации процессов, способствующих выработке и поступлению в кровяное русло плода веществ, необходимых для осуществления органогенеза. Возникает необходимость в активной синтетической деятельности хориального эпителия, который в то же время должен обеспечить увеличение своей массы за счет процессов пролиферации. Сочетание интенсивной пролиферации и синтеза специфических продуктов в период плацентации является особенностью хориального эпителия, в котором цитотрофобласт играет роль камбиального слоя.
Исключение составляют клетки цитотрофобласта с полиплоидным количеством ДНК, не трансформирующиеся в синцитий и сохраняющиеся в течение всей беременности. Возможно, именно эти клетки участвуют в синтезе хориального гонадотропина. Возникновение полиплоидии в части элементов цитотрофобласта можно рассматривать в качестве компенсаторной реакции, направленной на сохранение метаболической активности. В ядрах синцитиотрофобласта при нормальном развитии не происходит редупликации ДНК. Все энергетические ресурсы синцития, о чем свидетельствует высокая ферментативная активность, направлены на синтез специфических белков.
Таким образом, в незрелой плаценте компенсаторно-приспособительные реакции при физиологической беременности проявляются на клеточном и субклеточном уровнях в виде пролиферации, полиплоидизации цитотрофобласта и высокой синтетической активности синцитиотрофобласта. Учитывая транспортную роль клеток Кащенко—Гофбауэра, эти элементы иногда связывают с компенсаторными реакциями в ранние сроки беременности.
Во второй половине беременности в зрелой плаценте, несмотря на выключение части ворсин из обменных процессов в связи с местными дистрофическими и циркуляторными нарушениями, общая обменная площадь увеличивается. Это обусловлено рядом компенсаторно-приспособительных процессов, поддерживающих жизнедеятельность плаценты вплоть до родов. Формированию площади, достаточной для насыщения крови плода кислородом, способствует рост резорбционных ворсин. Несмотря на значительное отставание массы плаценты от массы плода во второй половине беременности, к родам на 1 кг массы тела ребенка приходится 4—4,5 см3 площади ворсинчатого хориона, что превышает площадь альвеол легкого на 1 кг массы тела взрослого человека. Темп дифференцировки ворсин оказывается максимальным к началу 28-й недели беременности, когда значительно возрастает количество мелких ворсин диаметром 10—90 мкм.
Увеличение количества резорбционных (молодых) ворсин связывают не только с ростом терминальных ворсин, но и с отпочковыванием «юных» ветвей от стволовых. Образование «юных» ворсин принято относить к числу компенсаторных реакций. Мы полагаем, что в качестве компенсаторных элементов следует учитывать лишь терминальные ворсины, имеющие сформированные синцитиокапиллярные мембраны. «Юные» ворсины, которые содержат слой цитотрофобласта и слабо развитую капиллярную сеть, удаленную от трофобласта, не в состоянии осуществлять компенсаторные реакции, поэтому их можно рассматривать в качестве резервных элементов [Калашникова Е. П., 1975]. Росту терминальных ворсин сопутствует развитие капиллярной сети с периферическим расположением капилляров, образованием синусоидов и синцитиокапиллярных мембран.
Образование синцитиокапиллярных мембран, через которые непосредственно происходят транспорт и газообмен, является важнейшим компенсаторным механизмом второй половины беременности. Формированию мембран способствуют «исчезновение» клеток Лангханса, образование безъядерных зон синцития и приближение капилляров к мембране трофобласта.
К числу компенсаторно-приспособительных реакций зрелой плаценты можно отнести образование синцитиальных узелков, обладающих высокой функциональной активностью. Снижение активной функции этих структур (исчезновение ШИК-положительных и базофильных гранул, активности щелочной фосфатазы, СДГ, пикноз ядер) сопровождается и уменьшением выраженности компенсаторных реакций.
К реакциям, направленным на поддержание функции фетоплацентарного комплекса, относятся также особенности плацентарного кровообращения. Прежде всего необходимо иметь в виду мощную сеть спиральных артерий, устья которых лишены способности сокращаться и расширяться. Большая роль принадлежит саморегуляции маточно-плацентарного кровообращения. Этому процессу способствует наличие миометральных и плацентарных артериовенозных шунтов. Компенсация кровообращения обеспечивается наличием венозных устьев в сети капилляров ворсин хориона, которые создают артериовенозные анастомозы. Последние вместе с периваскулярной капиллярной сетью являются буферной системой плаценты. Значительная емкость сети капилляров, превышающая емкость артериальных сосудов плодовой части плаценты, создает дополнительный бассейн крови.
Таким образом, во второй половине беременности основными компенсаторными механизмами являются увеличение количества терминальных ворсин малого калибра с гиперплазией и периферическим расположением капилляров, уменьшение толщины плацентарного барьера с истончением синцития, образование синцитиокапиллярных мембран и синцитиальных узелков, а также особенности плацентарного кровообращения.
В целом компенсаторно-приспособительные реакции плаценты можно представить как сложный динамический процесс адаптации элементов плацентарного барьера, включающих изменения на субклеточном, клеточном и тканевом уровнях, направленный на поддержание гомеостаза фетоплацентарной системы в соответствии с потребностями плода.
Развитие компенсаторно-приспособительных реакций в физиологических условиях происходит постепенно, по мере созревания плаценты. При воздействии патологических факторов наибольшей подвижностью обладают гемодинамические приспособительные реакции, морфологическим эквивалентом которых служит изменение площади сечения сосудов. Гемодинамические приспособительные механизмы реагируют в первую очередь на колебания изменившихся условий внешней среды и прежде всего на содержание кислорода в материнской крови, а также на изменения гомеостаза в организме матери и плода. Как показали проведенные нами экспериментальные исследования [Федорова М. В., Сапелкина И. М., 1979], под влиянием патологического ацидоза в крови беременных кроликов и их плодов происходят нарушения плацентарного кровообращения. Компенсаторно увеличивается число функционирующих капилляров, расширяются посткапилляры и венулы ворсин хориона. После лечения, направленного на устранение патологического ацидоза, наблюдается нормализация просвета сосудов. Т. Tominaga и Е. Page (1966) обнаружили расширение просвета сосудов хориона при острой гипоксии плода и сужение при повышении уровня кислорода в среде, омывающей ворсины хориона.
При хронической гипоксии возникают реакции, для осуществления которых требуется длительное время; при этом нарастает количество ворсин небольшого диаметра, уменьшается толщина эпителия хориона. В эксперименте при длительной гипоксии плода выявляется увеличение количества функционирующих ворсин и размеров синцитиокапиллярных мембран. При анемии у беременных обнаруживается уменьшение толщины синцития и увеличение числа ворсин с большим количеством капилляров [Степанова О. М., 1970]. При гипоксии плода компенсаторной реакцией является увеличение количества митохондрий и рибосом в синцитиотрофобласте. Повышается активность щелочной фосфатазы и ферментов, участвующих в анаэробных процессах.
Морфологический субстрат компенсаторно-приспособительных реакций в норме и патологии сходен. Наибольшей интенсивности компенсаторные реакции достигают при физиологической беременности. При хронической недостаточности плаценты уровень их в плаценте в целом понижен, хотя в периферических отделах некоторые показатели оказываются даже более высокими, чем в норме. По-видимому, периферическая часть плаценты является своеобразной резервной зоной для осуществления компенсаторных реакций, как было показано на примере поздних токсикозов беременных [Войтенко Л. М., Калашникова Е. П., 1976].
Одной из главных причин расстройства компенсаторных механизмов является нарушение созревания плаценты. Незрелость структурных элементов затрудняет адекватную компенсаторную реакцию плацентарного барьера и служит основанием для срыва адаптационных механизмов фетоплацентарной системы.
Рассматривая морфологические признаки компенсаторных механизмов при различных формах патологии, следует различать процессы, направленные на биологическую сохраненность тканей плаценты, и реакции, поддерживающие нормальную функцию всего фетоплацентарного комплекса. Первые реакции не всегда способствуют адаптации целостной системы мать—плацента— плод, так как могут сопровождаться нарушением маточно-плацентарного кровообращения, изменениями проницаемости мембран и т. д. Например, отложение фибриноида при физиологической беременности служит средством иммунной защиты и относится к приспособительным механизмам.
В то же время избыточное отложение фибриноида в межворсинчатом пространстве в условиях патологии нарушает циркуляцию материнской крови, и этот процесс нельзя рассматривать в качестве компенсаторной реакции фетоплацентарной системы. При физиологической беременности нарастание количества мелких ворсин в плотной части котиледона способствует увеличению площади обмена. В патологических условиях появление большого количества резорбционных ворсин, тесно прилегающих друг к другу, в центральной части котиледона затрудняет поступление материнской крови в периферическую часть котиледона и, таким образом, не может служить компенсаторным процессом в пределах фетоплацентарной системы.
Следовательно, изучение приспособительных реакций плаценты, как и других органов, указывает на то, что защитно-приспособительные процессы могут быть правильно оценены только в динамике с учетом характера, степени тяжести и длительности течения основного патологического процесса.